Conservation Status of the
Barndoor Skate (Raja laevis)
prepared by
Trevor J.
Kenchington Ph.D.
Gadus Associates,
Musquodoboit Harbour, Nova Scotia
for
Trawlers Survival Fund
August 1999

1 Introduction
Last year, in a short and controversial paper, Casey and Myers (1998a) reviewed the conservation status of the barndoor skate (Raja laevis) and concluded that it is “close to extinction”. They further suggested that it “could become the first well-documented example of extinction in a marine fish species”, while they also implied that the demise of the barndoor had resulted from these skates being taken as a bycatch in fisheries for other species. Finally, it was suggested that the advent of directed fisheries for other skates, following the collapse of more traditional fisheries, could result in extinction, while the closure of extensive areas to all trawling was proposed as “the only hope for the long-term survival” of barndoor (Casey & Myers 1998a).
This suggestion that an open-sea species could be driven to biological extinction by fishing, and indeed by fishing directed towards other species, was a dramatic departure from conventional wisdom. It also ran counter to long experience in the ocean, which has shown that the worst mankind can do has yet to eradicate entirely even species highly vulnerable to exploitation, such as the right whale. While freshwater, diadromous, coastal and reef species can be (and have been) driven to extinction by assorted human pressures, and some oceanic marine mammals may yet be, a fish such as barndoor had seemed invulnerable. Casey and Myers, (1998a) suggestion that to the contrary was, therefore, a radical idea, deserving of close scrutiny.
In 1999, a petition was made to the National Marine Fisheries Service (NMFS) to have the barndoor skate listed under the Endangered Species Act. Casey and Myers’ (1998a) paper was advanced as the primary scientific support for such a listing. In connection with NMFS review of that petition, a consortium of fishing-industry interests asked Gadus Associates to examine the issue. This document represents the initial findings of that examination. It has been prepared under a tight deadline, during a period when many government scientists were away from their offices. As a result, I have not been able to access all of the existing and relevant information which should be considered. Thus, in some of what follows I sometimes can only suggest points that merit further consideration by NMFS during its own, less hasty, review process. This document is largely confined to a review of the case for the “near extinction” of barndoor stake advanced by Casey and Myers (1998a). I do not attempt to discuss the legal or policy aspects of its proposed listing under the Endangered Species Act, though I do provide a few comments that may aid such discussions.
It should be understood that there is very little scientific knowledge of the barndoor and little enough of any skate. Early studies, up to the 1950s, served to document the species’ physical characteristics and its distribution, while suggesting its probable taxonomic relationships with the other skates. A scatter of incidental observations has provided hints of its abundance, food preferences, parasite burden and habitat requirements – information summarized by Bigelow and Schroeder (1953a) and Scott and Scott (1988). However, the last attempt at a biological study of this species was published a quarter-century ago by McEachran & Musick (1975), as part of a wider examination of northwest Atlantic skates, and even they had too few records of barndoor to say much about it. Clearly, there is a great deal that remains unknown.
2 Biology of Barndoor Skate
The barndoor is the largest skate in the northwest Atlantic, growing to about 1.5 m in length and 20 kg in weight[1]. It is one of a group of closely-similar species which includes Raja batis in European waters and R. floridana off the southern United States (Bigelow & Schroeder 1953b)[2]. Its reported range extends from as far north as southwestern Grand Bank and the southern Gulf of St. Lawrence, south to waters off northeastern Florida (Scott & Scott 1988) – though McEachran and Musick (1975) suggested that the most southerly records may have been misidentifications of R. floridana and that R. laevis may not occur south of Cape Hatteras. Kulka et al. (1996) have recently published charts of the distribution of “barndoor” as reported by observers aboard commercial fishing vessels working off Newfoundland. Those show the species as occurring in deep water off central Labrador, north to about 57°N latitude. That, however, is such a dramatic extension to the previously-reported range as to cast doubt on the observers’ specific identifications of the often-awkward skates.
Within this broad geographic range, barndoor have been found from the tideline (and even stranded above it: Bigelow & Schroeder 1953a) to depths of 430 m[3], though Kulka et al.’s (1996) charts suggest that they may be abundant at substantially greater depths. Bigelow and Schroeder (1953b) claimed that the preferred depth range was 10 to 140 m but did not specify any supporting evidence. In contrast, Casey and Myers (1998a), in an aside in a footnote, stated that barndoor were “commonly found” at 200 to 400 m depth (with the implication that they were more common there than at lesser depths) on the slopes of St. Pierre Bank “historically” – by which they appear to have meant in the 1950s and 1960s. Thus it is not presently clear whether the species was primarily one of the banks or the continental slope, in the era before it was markedly impacted by fishing. It does seem more of a deepwater species now.
Barndoor skate appear to tolerate temperatures from barely above freezing to 20°C or more, as well as salinities from ocean water down to about 20‰ and maybe even below. In the Mid-Atlantic Bight, they may move away from the beach to avoid the warm shallows in the hottest months of the year, while in the north they may move off in winter to escape extreme cold (Bigelow & Schroeder 1953b).
In shallower areas, barndoor are normally taken on sand or gravel bottoms but they are found on mud at greater depths (Bigelow & Schroeder 1953b). It has been suggested (though on questionable grounds) that they are less inclined to hug the bottom than other skates are and it is certain that they are strong swimmers (Bigelow & Schroeder 1953a). They have wide-ranging tastes, eating mostly lobsters, crabs and shrimps but also isopod crustaceans, bivalve and gastropod molluscs, squids and worms. They also eat a range of fish, including some commercial types. In European waters, Raja batis is known to eat cod and the same may be true of barndoor (Bigelow & Schroeder 1953a,b), though presumably only small juveniles could be taken.
Nothing is known of their predators (Scott & Scott 1988). Little except large sharks could tackle an adult barndoor but the juveniles may be vulnerable to a wider range of large fish and perhaps also to seals. A range of parasites has been recorded (Scott & Scott 1988).
Like the other skates, barndoor spawn a small number of large eggs (very large in the case of barndoor: around 130 mm long). Those have been recorded on both stony and silty bottom (Vladykov 1936). From the very limited information available, barndoor seem to spawned during the winter at all depths and in all areas inhabited by the species. The young apparently hatch in late spring or early summer, at a length of about 180 mm (Bigelow & Schroeder 1953b).
No useful information is available on the population dynamics of barndoor – no growth rates, fecundity rates, mortality rates or the like (Scott & Scott 1988). Raja batis, a much larger animal (reaching over 2.5 m in length and 50 kg in weight), is thought to become sexually mature at 11 years of age (Brander 1981) but it is far from sure whether the same applies to R. laevis. Indeed, the ageing of skates and other cartilaginous fish is sufficiently problematical that Brander’s (1981) figure should not be unconditionally accepted even for the European species.
Interestingly, barndoor is the only northwest Atlantic skate with an extended history of exploitation. Goode (1884) noted them (and the other skates) primarily as “troublesome to the fisherman, clogging his lines and pound-nets” but did record that barndoor were sometimes salted by New Hampshire fishermen (though not why – perhaps it was for use as lobster bait) and that a few New York restaurants had begun serving barndoor “wings”, as Europeans had long eaten Raja batis. By the 1950s, Bigelow and Schroeder (1953b) noted barndoor (with a few R. ocellata) as the primary species in the very limited New England skate landings. Only the wings (which they called “saddles”) were marketed. Leim and Scott (1966) and Scott and Scott (1988), however, saw barndoor as essentially unused in Canadian waters. Only the incidental catch taken in weirs was sometimes landed, and then only for processing into pet food and fish meal.
The new skate fisheries of the 1990s, which are being developed in the wake of the loss of traditional groundfish resources, are targeted on other, smaller species. Barndoor are, presumably, now too scarce to have attracted the interests of potential developers, at least in those, more northerly, areas where the loss of groundfish has severely affected the fishing industry.
3 Depletion, not Extinction
While I argue below that there is no evidence that the barndoor skate is in danger of biological extinction, and indeed that there is persuasive evidence that it is not in any such condition, that should not be misinterpreted as an argument that the barndoor is near its natural abundance.
Large flatfish are, by their nature, vulnerable to depletion by trawl fisheries targeted on smaller flatfish or almost any size of roundfish. This is a straightforward and well-understood consequence of the size-selective properties of trawl nets[4], which retain even quite small flatfish. Species with large adult sizes are retained (and therefore commonly killed) at a relatively early stage in their lives, resulting in more pronounced depletion than most bycatch species experience. Larger species are also, typically, longer lived, with lower natural death rates (technically “Natural Mortality rates”) and so an increased susceptibility to depletion under any given level of fishing effort (technically equivalent to the “Fishing Mortality rate”).
Like sharks and dogfishes, skates are particularly vulnerable to depletion under any increase in their death rates, as a result of their low fecundity. Where the bony fish typically place their reproductive energies into spawning very large numbers of eggs (such that environmentally-controlled survival of their offspring largely determines the abundance of the next generation), the cartilaginous fish are either live-bearers or spawn a few, large eggs (as does the barndoor), which have the advantage of being less susceptible to the many dangers of the marine environment. When an additional cause of death, such as fishing, is introduced, however, these fish have only a limited ability to compensate. Their populations typically decline sharply under directed exploitation. Even the (usually lesser) pressures on a bycatch species caused by incidental capture in fisheries targeted on other resources could lead to depletion, though ecosystem complications can sometimes lead to increases in such species, as was seen with the smaller skates on Georges Bank in the 1980s (Fogarty & Murawski 1998).
Barndoor, being both a very large flatfish and a low-fecundity skate, should therefore be expected to have declined in abundance once it began to be taken as a bycatch in the cod fisheries, beginning some 500 years ago. That decline should have accelerated once trawl fisheries were developed in the northwest Atlantic (something which began early in the 20th Century but expanded greatly in the years after 1950), though the degree of the barndoor’s depletion will always have been limited by the absence of any directed fishing for the species.
There is scattered evidence that such a decline did indeed occur. As discussed below, there are hints of it in research-vessel survey data, particularly from St. Pierre Bank in the 1950s and from U.S. waters in the mid- and late-1960s. Less-quantitative indications of earlier depletion can be drawn from some published accounts. Bigelow and Schroeder (1953a), for example, described barndoor as “a common fish in all parts” of the Gulf of Maine and “plentiful” off the Atlantic coast of Nova Scotia, while they noted that it was “often stranded on the beach” as a result of straying into very shallow water. They reported on one trip by a commercial trawler in 1951, working 42 hauls from the Nantucket Lightship to central Georges Bank, which took an average of 21 barndoor per haul. Elsewhere, those authors reported a trip to Georges in 1929, which took 42 barndoor in 37 hauls, and two (undated) trips off southern New England which together took 441 in 107 hauls (Bigelow & Schroeder 1953b)[5]. Most interesting of all was an August to October 1943 effort to monitor the catches (including bycatches) by a flounder trawler out of Stonnington, Connecticut, which worked an 80 ft trawl (Merriman & Warfel 1944). The average barndoor catch was 25.73 lb. per hour of fishing. Assuming that the net had an effective wingspread of about 20 m and was towed at about 3 knots, that indicates a catchable biomass (not corrected for the efficiency of the gear) of about 80 kg of barndoor per square kilometre on the flounder grounds[6]. While these assorted numbers are not directly comparable with modern survey-based estimates of skate biomass, they do point to a level of barndoor catches far greater than anything normally experienced in the 1990s – and hence to the depletion of the species.
While such depletion is only to be expected and seems certain to have occurred, it should not be confused with an approach to biological extinction – an entirely different state. Instead, there is every reason to suppose that the barndoor population long-since stabilized at a low level in approximate balance with the fishing pressures acting on it and even (as discussed below) that it may recently have increased somewhat. Rather than a one-way trip to oblivion for the species, this depletion represents but one facet of the conversion of the northwest Atlantic from a natural ecosystem into an exploited one – a conversion analogous to that which distinguishes a sustainably-managed wood lot from a virgin forest and, more distantly, a wheat field from natural prairie grassland. While its inevitability may be regretted, such depletion (to a point far short of extinction) is an unavoidable and expected consequence of the growth of the Atlantic fisheries.
4 Casey & Myers’ Argument
In developing their argument that the barndoor is in danger of extinction, Casey and Myers (1998a) made a number of points. They noted the general vulnerability of skates to over-fishing (as above) and the increased abundance of some smaller skate species on Georges Bank in the 1980s (apparently as their biomass replaced that of depleted commercial groundfish), followed by the decline of those fish when a directed fishery developed (cf. Fogarty & Murawski 1998). Of more immediate relevance to their conclusion that the barndoor is near extinction, Casey and Myers (1998a) argued:
· Barndoor were once common.
· Estimates of biomass density, from research-vessel surveys, showed a decline in their biomass into the early 1970s, after which barndoor were only taken by the survey vessels on Browns and Georges banks. (On St. Pierre Bank, Casey and Myers, (1998a) estimated barndoor populations fell from some 600,000 individuals in the 1950s to less than 500 in the 1970s.)
· Barndoor skate are vulnerable to commercial trawl nets (of 70 to 140 mm mesh) at all ages from hatching onwards.
· The mortality rate required to drive the barndoor to extinction is about 0.4 on Georges Bank and closer to 0.2 in the colder water further north.
· Fishing mortality rates imposed on cod have long been above 0.4 and often much greater.
· The closely-similar Raja batis is now locally extinct in the Irish Sea.
Casey and Myers (1998a) also suggested that areas at great depth “may have represented one of the last refuges” of the barndoor, while the species has survived on Browns and Georges banks “probably because of a faster growth rate combined with the seasonal closure of parts of these banks to trawling”.
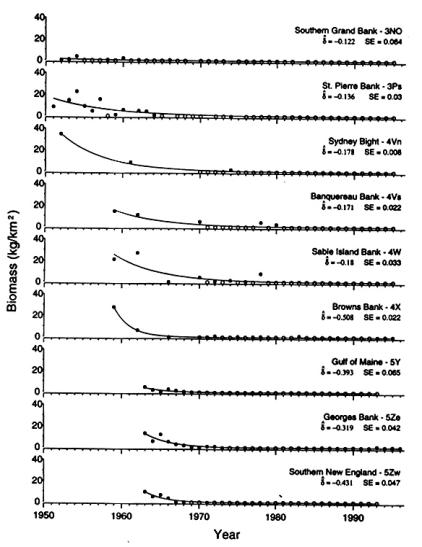
Casey and Myers’ estimates of biomass density
(after
Casey & Myers 1998)
The starting point, that barndoor were once common and no longer are so, is fully justified but does not amount to a case for the species being anywhere near the verge of biological extinction (see above). The remainder of Casey and Myers’ (1998a) argument does not stand up to scrutiny[7].
5 A Rebuttal
5.1 Trends in Barndoor Biomass
Aside from repeating some of the pre-1960 published observations of barndoor abundance that are outlined above, Casey and Myers’ (1998a) evidence for a decline in the biomass of the species was based entirely on data from research-vessel groundfish surveys conducted in various areas from southern Grand Bank to the area around the Nantucket Lightship (which they misleadingly termed “southern New England”). Notably, they did not include any information from the rest of the Mid-Atlantic Bight, nor any from the Gulf of St. Lawrence. Since the surveys from which the data were drawn were primarily designed to support assessments of the major continental-shelf resources, particularly cod and haddock, none have quite covered the full depth range of the barndoor, though the more recent ones have extended down to 365 or 400 m. Finally, as presented by Casey and Myers (1998a), the various survey time series ended in 1993 to 1996, depending on the area concerned. Thus, their conclusions took no regard of events in the last three to six years
For those areas which were covered by the surveys that they used, Casey and Myers (1998a) claimed that the “population trend is similar for all regions”. That was a questionable reading of the data they presented, which deserve to be considered individually. Although they nowhere explicitly said as much, their conclusion of “near extinction” implies that the supposed biomass decline is on-going. That conclusion is not supported by the research-vessel data.
5.1.1 St. Pierre Bank
For St. Pierre Bank (NAFO Division 3Ps), Casey and Myers (1998a) showed barndoor biomass densities of about 10 to 25 kg/km2 in the early 1950s, which fell to below 20 kg/km2 in the second half of that decade and then to below 10 kg/km2 in the early 1960s[8]. Since 1964, barndoor have only been taken in the routine surveys on the Bank once – in 1974. On its face, these densities appear to show that the barndoor biomass on St. Pierre Bank was reduced by some 75% through the 1950s and that the species was all but extirpated from the area by 1965. In reality, the data cannot support such an extreme conclusion.
Research-vessel surveys began on St. Pierre Bank in 1950 and they have continued to date. However, there has only been a continuous series, suited to long-term monitoring of trends in biomass, since 1972 (Pitt et al. 1981). Even that series was interrupted around 1980 by a change in the survey vessel and gear from the side trawler A.T. Cameron, which worked a Yankee 36 trawl, to the stern trawler Wilfred Templeman and her Western IIA net.
From 1972 onwards, the surveys have used a stratified-random sampling design, suited to the estimation of biomasses across the Bank and down to depths of 400 fathoms (731 m) around it, though the surveys ignored a wide swath of coastal waters, including Hermitage and Placentia bays (Pitt et al. 1981). Within limits (discussed below in connection with the Scotian Shelf surveys), this series can be used to document the long-term trends in barndoor biomass over the last quarter century but, since essentially none of the species have been caught in that time, the data are not very informative.
There is little published documentation on the earlier surveys, conducted before 1972, though it is known that they began as exploratory surveys, designed to find concentrations of haddock for the commercial fleet, with trawling stations being arrayed along transects that were selected in an ad hoc manner. Through the later 1950s and into the 1960s, these transects gradually became fixed as the intention of the work turned to the estimation of biomass. As finally established, there were eight transects in Division 3Ps (Pitt et al. 1981). While those provided a consistent picture of biomass trends (primarily in the shallows, where haddock were expected) for about a decade, the data from them cannot readily be compared to later biomass estimates. Casey and Myers (1998a) attempted to do so by allocating each survey tow to the modern stratum in which it fell and scaling up from the catch per tow to the biomass in the stratum, just as data from a stratified-random survey would be scaled. The old transects were not, however, randomly placed within the strata. Thus, Casey and Myers’ (1998a) approach cannot fail to have produced biased results (though whether biased upwards or downwards is not known). Moreover, of the 30 or more modern strata in Division 3Ps, only about half contained any of the old transect stations. It is not clear how Casey and Myers’ (1998a) estimated biomasses for the others but further bias was inevitable.
The A.T. Cameron came into service around 1960 but I am not presently aware of exactly when she took over the St. Pierre bank surveys. Before she did, they were conducted by much smaller side trawlers, working different gear – though no detailed records of the nets used are available to me[9]. It is not clear how Casey and Myers (1998a) corrected for this change in trawls or even if they did. Since no comparative-fishing experiments were conducted, that correction (or its absence) must have introduced yet another bias, of unknown direction and magnitude.
The possible size of this latter bias should not be under-estimated. In the late 1960s and early 1970s, Soviet research ships worked off the U.S. east coast, supplementing American efforts at biomass surveys of what was then an international resource. A number of comparative-fishing trials were run in order to inter-calibrate these different surveys. One interesting observation was that the Soviet side trawler Albatros, working a net with a simple groundrope, caught about six to ten times the weight of skates (all species combined[10] and depending how the figures were calculated) than did the U.S. stern trawler Albatross IV, which fished a net with bobbin gear. The two nets had similar wingspread and exchanges of them between the ships showed that the vessel effect, while present, was minor. Hence the difference in catchability related to the efficiency of the gear (Hennemuth 1968; Grosslein 1968). A later trial found that the standard Yankee 36 net (used in the U.S. surveys from 1963 onwards) took only one twelfth the catch of skates as the Soviet 27.1 net, despite the two again having similar wingspreads (Griswold & Efanov 1972). It is supposed that these differences arose because bobbin gear tends to roll over skates which would be caught by a groundrope which tended the bottom more closely[11].
Since Casey and Myers (1998a) did not indicate that they had considered correcting their biomass estimates for such pronounced differences in the efficiency of different survey gears, the possibility remains that their figures for the years before the A.T. Cameron[12] took over the surveys may be biased upwards by an order of magnitude – if the vessels used for the surveys of the 1950s towed nets with simple groundropes, rather than bobbin gear.
Thus, what the Division 3Ps data really show is that early surveys, partly directed for haddock, using an unknown small net, yielded catches from which Casey and Myers (1998a) calculated (by undocumented means) an average barndoor biomass density of the order of about 15 kg/km2. As the transect design was progressively changed from a haddock-directed one into a fixed, representative design, equivalent calculations led to a density of about 10 kg/km2. When the A.T. Cameron took over the surveys, she may have had a year or two of barndoor catches (or perhaps those were the last years under previous vessels) but thereafter took essentially none on St. Pierre Bank. Whether any of the apparent biomass trend through those years was actually linked to the abundance of barndoor skate, or whether it was all an artifact of survey gear or protocols, is a matter for speculation. The latter is by no means improbable.
That being said, it remains possible that the skates did decline from 1950 to 1965, even if that trend was not detected in any reliable way. If so, the facile assumption that the change was a result of fishing pressure should not be uncritically accepted. Barndoor is a bottom-feeding fish that reached the northerly limit of its range on St. Pierre and Grand banks. In that much, it was similar to the much better studied haddock and the history of the latter is indicative of possible trends in the skate.
In the 1920s, one of the largest haddock fisheries in Canada was at Ingonish (at the northern tip of Cape Breton), where it exploited migrants entering the Gulf of St. Lawrence(Needler 1931). Those fish occupied extensive areas of the southern Gulf in the summer, as they had for many years (e.g. Bell 1859; Cornish 1912; Cox 1921). A few were even known to stray as far north as the Strait of Belle Isle, between Newfoundland and Labrador (Huntsman et al. 1954). By 1940, however, although there were still haddock fisheries on St. Pierre Bank and in Sydney Bight, the centre of the fishing had shifted markedly to the westwards (McKenzie 1946). Haddock remained in the Gulf, seasonally, but apparently no longer in such abundance as they once had (e.g. McKenzie 1959). Through the 1950s, they continued to support catches in St. Georges Bay, in the southeastern corner of the Gulf (Scott 1977) but Kenchington (1980) found no sign of the species there during extensive surveys in 1977. By 1990, by commercial standards, they had disappeared from north of a line from roughly Halifax to Sable Island, 300 km from the once-rich fishing ground off Ingonish (Kenchington & Halliday 1994; Kenchington 1996), though isolated specimens were still sometimes seen.
While excessive fishing pressure has been blamed for the final eradication of haddock, as commercially-viable resources, from St. Pierre and Grand banks, it is hard not to conclude that their general withdrawal from the vast area from the mouth of the St. Lawrence to eastern Grand Bank over a period of decades was not ultimately caused by some unknown shift in the marine environment[13]. While there can be no firm evidence, it is entirely possible that the barndoor skate has been influenced by the same shift as affected the haddock, potentially reducing its abundance on St. Pierre Bank by a substantial fraction.
Thus, contrary to Casey and Myers’ (1998a) claim, there is no solid evidence for any change in the biomass density of barndoor skate on St. Pierre Bank. There is no evidence whatsoever of a continued reduction in barndoor biomass in the area after 1965. If there was a pronounced decline through the 1950s and into the 1960s, which remains possible, there is no compelling reason to suppose that it was caused by fishing alone, though that is one plausible cause. Rather, the area seems to have undergone a long-term climatic shift which would have been consistent with the claimed decline.
5.1.2 Grand Bank
All that has been said above of the barndoor-biomass data from St. Pierre Bank is also true, to varying degrees, of the data from Grand Bank (NAFO Divisions 3NO). In addition, as befits the extreme edge of the range of the species, its estimated biomass densities on that Bank were always very low. Even in the early 1950s, Casey and Myers’ (1998a) calculated densities never reached 10 kg/km2 and once the A.T. Cameron took over the surveys, no barndoor were taken at all, bar tiny amounts (perhaps one individual each year) in 1960, 1962 and 1966. Not a single example of the species has been caught in the surveys since they took on their modern, stratified-random form – which, in Divisions 3NO, was in 1971 (Pitt et al. 1981). Clearly, such limited data cannot even indicate, let alone prove, anything of the current conservation status of barndoor or of past trends in its abundance.
For this area (though also with reference to Division 3Ps), an additional relevant body of data appeared shortly before Casey and Myers’ (1998a) paper, though they did not cite the report and may not have been aware of it. In 1996, Kulka et al. (1996) had summarized available information on the skates of the Grand Banks, primarily with reference to the thorny skate (Raja radiata) but also covering each of the other species which occurs there. They looked at both research-vessel catch rates from 1991 to 1995 (part of the same data set that Casey and Myers used) and at data collected by observers aboard commercial vessels from 1981 to 1994. Both data sets were presented as catch rates averaged over five-year periods, with local averages being calculated for each small area for which at data had been collected.
As Casey and Myers (1998a) had shown, there were few barndoor in the research vessel catches and Kulka et al. (1996) did not mention the species in conjunction with that data set. The observers monitored a large number of commercial catches in which “barndoor” comprised, on average, 4% of the skate caught (which together comprised 1% of the total catch). They were found primarily in deeper waters around the edge of the banks, particularly along the continental slope. In 1981-85, “barndoor” were seen in bycatches all along the continental slope from off Labrador (about 57°N latitude) south to the vicinity of Funk Island Bank, around Flemish Cap, on the southwestern edge of Grand Bank and also in the Laurentian Channel west of St. Pierre Bank. In 1986-90 their apparent range was more contracted, with few being taken other than along the seaward edges of Funk Island Bank, southwestern Grand Bank and St. Pierre Bank. By 1991-94, few were seen other than off Funk Island. Where they did occur, however, the five-year averages of catch rates almost always exceeded 1 kg per hour of fishing. It is not clear to what extent the apparent distributional changes resulted from real trends in barndoor abundance over the years, as distinct from changes in target species, gear and fishing techniques in fisheries for which barndoor is only a very minor bycatch. Thus, there are limits to what can be drawn from this dataset.
Moreover and as noted above, in view of the massive northern extension to the reported range of the species, there must be some doubt as to whether the “barndoor” recorded by the observers really were all Raja laevis. This might be resolved by discussions with the managers of the observer program and with a selection of the observers themselves. Until it can be, the data cannot be unconditionally accepted.
To the extent that they are genuine, they certainly throw Casey and Myers’ (1998a) conclusions into question. Barndoor clearly were not taken in the research-vessel surveys on Grand Bank after 1966 but, unless the observers were in error, those fish were present in not-inconsiderable numbers throughout the 1980s, well into the 1990s and perhaps up to date. Clearly there is a pressing need to make a more detailed analysis of the data to explain this disagreement. It might be a matter of the fishing gear used, since the observers made some trips on longliners and gillnetters, as well as trawlers. It might be a question of the depth of the fishing, some commercial trips having operated at greater depths than the research vessels routinely do (Kulka et al. 1996). Such an analysis should also consider whether or not the observer data show a decline in “barndoor” catches from 1981 to date, indicative of on-going depletion of the species.
Until such careful consideration has been completed, it would be premature to reject, or to fully accept, either Casey and Myers’ (1998a) or Kulka et al.’s (1996) representation of the abundance of barndoor on Grand Bank[14].
5.1.3 Gulf of St. Lawrence
Casey and Myers (1998a) made no mention of the Gulf of St. Lawrence, an area where barndoor skate were once reasonably abundant. Much as other parts of Canadian waters, the southern Gulf (NAFO Division 4T) has seen a mix of groundfish surveys from 1951 to 1970, followed by a regular series from 1971 onwards (Halliday & Koeller 1981). There seems no reason why the area should not be included in any assessment of the status of barndoor.
5.1.4 Sydney Bight and Eastern Scotian Shelf
In their plots of their biomass-density estimates, Casey and Myers (1998a) chose to present separately the data from Sydney Bight (NAFO Division 4Vn), the easternmost Scotian Shelf (Division 4Vs, which they misleadingly termed “Banquereau Bank”) and the central shelf (Division 4W, incorrectly termed “Sable Island Bank”). The three areas actually show very similar trends over time: a thin scatter of occasional surveys through the 1950s and 1960s, all but one of which produced biomass-density estimates for barndoor of between about 10 and 35 kg/km2, followed by a scatter of occasional catches through the 1970s and none at all after 1980. As with the surveys of St. Pierre Bank, however, these data may not be as conclusive as they at first appear.
Halliday and Koeller (1981) have summarized the survey cruises conducted in Divisions 4VW in the 1951-1969 period. Excluding a number of focused research cruises, fishery-development exercises and the like, they generated a list of those cruises which had the primary purpose of documenting groundfish distribution and abundance (Halliday and Koeller 1981, Table 1). Interestingly, their list bears little relation to the scatter of data points plotted by Casey and Myers (1998a) for those early years. The latter showed surveys in Division 4Vn in 1952 and 1961, while 1959 and 1962 had data for Division 4Vs. Halliday and Koeller (1981), in contrast, showed surveys in Division 4Vn in 1959, 1960, 1961, 1963, 1964, 1966 and 1969. For Division 4Vs, they noted surveys in 1959 and annually from 1962 onwards. For Division 4W, Casey and Myers (1998a) had data only for 1959, 1962 and 1966, whereas Halliday and Koeller (1981) listed surveys in 1951 and annually from 1958 onwards. Casey and Myers (1998a) may have had good reasons for using only a sub-set of the available data and for constructing biomass estimates using data from cruises that Halliday and Koeller (1981) did not consider as groundfish surveys. If so, those reasons have not been stated and, until they are, the estimated barndoor biomass-densities for Divisions 4VW in the years before 1970 must be regarded with suspicion[15].
Casey and Myers (1998a) also deliberately inflated their biomass estimates for the years before 1970 by applying a “correction” for the assumed difference in barndoor catchability between day and night. This was of no consequence to their data series from the waters off Newfoundland, where essentially no barndoor were reported by any surveys after 1966. However, from Sydney Bight southward, it served to increase the apparent severity of the depletion of the species. Casey and Myers (1998a) reasoned that the fish are more vulnerable to the nets by night than by day (when they are more likely to be pressed closely to the bottom), while the early surveys were conducted in daylight only. To calibrate those to the later data, the two authors “converted half of the catches before 1970 to night catches using a factor of 2.08” (Casey & Myers 1998a)[16]. It is not clear how that was done. The obvious approach would have been to scale all of the biomass estimates for years before 1970 by half the difference between the assumed day and night catchabilities – effectively increasing the early estimates by 154%. However, Casey and Myers (1998a) choice of wording suggests that they may actually have selected half the catches, multiplied the data by 2.08 and then re-calculated the biomass estimates. If so, the criteria by which certain catches were chosen for “promotion” to nocturnal ones would have the potential to introduce massive biases, particularly if the larger catches were those selected.
This whole “conversion” process was, in any case, suspect. It may well be true that the small vessels used for the earliest surveys worked only by day. However, the A.T. Cameron was suited to a two-watch, 24-hour system and certainly worked that way in the 1970s. Since she entered service by 1959 and undertook almost all of the survey cruises on the Scotian Shelf from 1961 onwards, Casey and Myers (1998a) “correction” did little more than unnecessarily inflate their biomass-density estimates for the 1960s, relative to those in subsequent years – thus exaggerating, if not actually creating, the impression that that barndoor declined on the Scotian Shelf between circa 1960 and 1970.
To these particular doubts must be added those already described in relation to the surveys on St. Pierre Bank concerning both the non-standard vessels and gear used (the A.T. Cameron not being employed in these surveys before 1959 nor invariably so before 1965) and the use of data from non-random stations in calculating pseudo-stratified biomass estimates. In short, while Casey and Myers (1998a) pointed to sharp declines in barndoor biomass in Sydney Bight and on the easternmost Scotian Shelf in the 1950s and 1960s, the data they used for that period are far too suspect to support any conclusions at all – at least until some more reliable analysis of them can be prepared.
From 1970 onwards, in contrast, the annual summer groundfish survey of the Scotian Shelf has provided a fairly consistent basis for biomass estimation. During those years, however, the catches of barndoor have been too poor for much to be deduced. The numbers of stations in Division 4Vn in these standard surveys are so few that it is hardly surprizing that barndoor have almost never been taken there – and only one has been, off Cape Smokey in 1974 (Simon & Comeau 1994; Casey & Myers 1998a). The other two Divisions saw a thin scatter of the fish, the only substantial catch being taken on the edge of the deep Sable Gully in 1978 (Simon & Comeau 1994; Casey & Myers 1998a).
The only pattern that emerges from these scant data is that all of the barndoor taken by the routine surveys on the eastern Scotian Shelf were caught from 1970 to 1980 inclusive, with none being taken thereafter. While that could be indicative of an on-going decline in the abundance of barndoor, it is also interesting that the trend coincides with the change in survey vessel from the side-trawling A.T. Cameron, with its Yankee 36 net, to first the Lady Hammond and then the Alfred Needler – both stern trawlers which each worked a Western IIA. The absence of barndoor in subsequent catches may therefore be a result of nothing more than a lower efficiency for that animal (which is only incidentally caught in surveys primarily targeted on commercial groundfish) with the new rig.
Comparative fishing trials were run to inter-calibrate the three vessels and so to maintain continuity of the surveys. However, barndoor were far too rare in the catches for such trials to indicate anything of the relative efficiency of ships and nets for that particular species. Such trials in any case tend to produce complex and confusing data which are hard to interpret in any useful way. It might, however, be worthwhile to review the trial data to see whether the Alfred Needler and her net are noticeably less efficient at catching other, smaller skates than the A.T. Cameron was. Until that is done, the possibility that the barndoor were not seen after 1980 simply because of an artifact of the survey method should be retained.
As in the case of St. Pierre Bank, even if the decline in barndoor biomass was real, it may have resulted (in whole or in part) from a slow climatic shift, rather than the effects of fishing. Divisions 4VW certainly saw a major range contraction of haddock through these same years.
5.1.5 Western Scotian Shelf and Bay of Fundy
Casey and Myers (1998a) presented another plot of estimated barndoor biomass-densities for an area which they described as “Browns Bank – 4X”. From the known distribution of catches in the 1970 to 1992 surveys[17] (Simon & Comeau 1994), it is certain that that plot actually relates to NAFO Division 4X, an area which extends from La Have Bank and the waters off Halifax, west to the Northeast Channel and thence north to encompass the entire Bay of Fundy. Browns Bank is but one minor component of this large Division.
Within it, Casey and Myers (1998a) presented data from three early surveys in 1959, 1962 and 1966, plus the regular Canadian July groundfish survey which has operated from 1970 to date. Those figures produced a very high biomass-density estimate for 1959, of around 30 kg/km2, a moderate one of about 5 kg/km2 in 1962 and, thereafter, a consistent pattern of intermittent, low estimates. Their plot provided no sign of a change in biomass in Division 4X since the mid-1960s.
As with the areas further to the northeast, it is far from clear either which early cruises Casey and Myers (1998a) chose to use data from or why they opted to reject others. The only cruise in 1959 in this area which Halliday and Koeller (1981) listed as a groundfish survey, for example, was a redfish survey along the edge of the continental shelf. From very recent experience (see below), that could be expected to have encountered relatively good catches of barndoor. Thus, it is entirely possible that the Casey and Myers’ (1998a) high estimate of biomass-density for 1959 was nothing more than an artifact resulting from the inappropriate use of this one limited survey as an indicator of barndoor densities throughout Division 4X. Since those authors made no attempt to explain how they derived their estimates, the validity of this sole indication that the species was ever markedly abundant on the western Scotian Shelf must remain in doubt.
For other early years, Halliday and Koeller (1981) listed survey cruises in Division 4X in 1952, 1963, 1966, 1968 and 1969. It is not clear why Casey and Myers (1998a) did not use all of the resulting data, nor where they obtained data for the Division from 1962. Moreover, The U.S. groundfish survey program, which commenced routine and standardized surveys in 1963, extended far into what are now Canadian waters from its conception (Azarovitz 1981). I can see no adequate justification for not including the time-series of biomass estimates from that work as part of the analysis of barndoor in Division 4X[18].
While Casey and Myers (1998a) plot of data from Division 4X follows those from the eastern Scotian Shelf in many ways, and shares many of the same imperfections, it does differ in one essential: It shows estimated barndoor biomass densities remaining constant (within the limited precision that these surveys can provide) through the 1970s, 1980s and the early 1990s. (As presented, the data series ends in 1995, even though the data from the 1996 and 1997 survey should have been available in time to be included in Casey and Myers’ paper.) That constancy may cast doubt on the possible lower efficiency of the Alfred Needler, relative to the A.T. Cameron, as a skate-survey platform which was suggested above – though it is, of course, possible that such a reduction in efficiency has merely masked an increasing barndoor biomass on the western Scotian Shelf. More importantly, the steady biomass estimates are persuasive evidence that the barndoor, after perhaps suffering marked depletion in earlier decades, has long since had a stable population. That is: The species is showing no sign of imminent extinction.
5.1.6 Georges Bank
For Georges Bank (NAFO Division 5Ze), Casey and Myers (1998a) relied on the U.S. groundfish surveys, which have been operated routinely in the fall of the year since 1963. They did not attempt to use any earlier data, such as they had done for the various Canadian areas, although there was a similar scatter of “pre-modern” survey cruises (Azarovitz 1981) on Georges[19]. Their resulting estimates of barndoor biomass density were around 10 kg/km2 in the mid-1960s but promptly fell to very low levels by 1969, where they remained consistently to the end of the data series used by Casey and Myers (1998a) in 1996. These estimates strongly indicate that the barndoor population on Georges Bank has been stable for thirty years (casting substantial doubt on any claim that the species is heading for extinction), though they also provide persuasive evidence of a sharp decline in abundance through the mid- and late 1960s.
While there have been some changes in ships and nets over the years, there was no obvious cause within the survey protocols which might have created that early decline as an artifact. Casey and Myers (1998a) do appear to have applied their procedure for “converting” some of the catches into “nocturnal” ones to the U.S. data from before 1970, just as they did to the Canadian equivalent, despite the U.S. surveys having involved 24-hour fishing ever since the series started in 1963. That will have exaggerated the rate of barndoor depletion but, unless there was a strange choice of which catches to “convert”, it will not have been sufficient to create a spurious decline in the biomass estimates during the 1960s. Thus, that decline must be accepted as genuine.
5.1.7 Gulf of Maine and Nantucket Lightship
The distinct (and unconnected) areas of NAFO Divisions 5Y and 5Zw were treated by Casey and Myers (1998a) in exactly the same way as they did Georges Bank. The resulting barndoor biomass-density estimates showed low levels of the skate in the Gulf of Maine in the mid-1960s but somewhat higher ones south of Cape Cod. Both had dropped to zero by 1970 and since then essentially no barndoor have been caught by the surveys in either area – at least, not by those surveys which Casey and Myers (1998a) used in preparing their plots.
Throughout the 1963 to 1993 period (when Casey and Myers ended their data series for these two areas, although they should have had access to data through to 1997), the Gulf and the waters immediately south of Cape Cod seem to have been less attractive to barndoor skate than Georges Bank was. Those areas appear to have experienced the same decline in the species through the 1960s as the Bank did, along with the same stability thereafter – albeit the population was “stable” at a very low level.
5.1.8 Mid-Atlantic Bight
Barndoor skate range at least as far south as Cape Hatteras and were once abundant even in Chesapeake Bay (Bigelow & Schroeder 1953b). From their inception in 1963, the regular U.S. groundfish surveys reached as far to the south westwards as to approach Hudson Canyon. In 1967, they were extended all the way to Hatteras and that area has been consistently surveyed ever since (Azarovitz 1981). Despite this, Casey and Myers (1998a) declared the barndoor skate to be “close to extinction” without examining any data from south or west of NAFO Division 5Zw. No such unsupported statement can be taken seriously.
5.1.9 Recent Trends
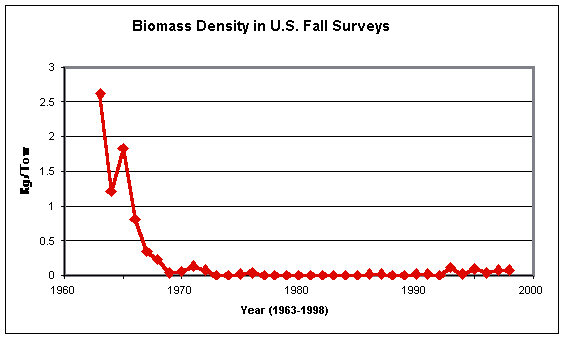
Casey and Myers (1998a) terminated their time-series plots at
various dates from 1993 to 1996, although all of the routine survey programs
that they relied on have been on-going. Data up to 1997 should have been
available in time for inclusion in their paper. At the time of writing (August
1999), the data from the 1998 surveys of all of the relevant series, along with
those from the U.S. spring 1999 surveys, should be available. Unfortunately,
the restricted deadline within which this document has had to be prepared
(coupled with key individuals being away from their offices) has prevented me
from compiling those data. Through the courtesy of Mr. Steve Murawski and
Ms. Kathy Sosebee at the Northeast Fisheries Science Center, however, a
simplified version of the results of the U.S. surveys can be shown:
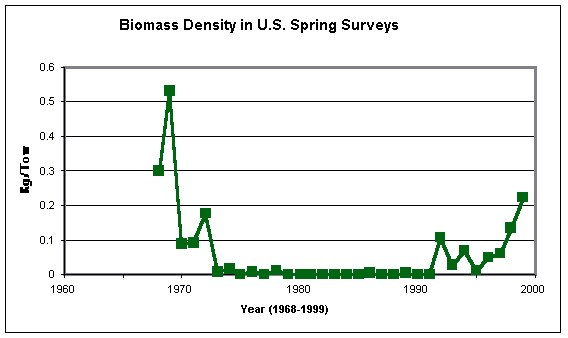
These figures present the data in a catch-per-tow, rather than catch-per-area-swept basis and thus are scaled differently to those shown by Casey and Myers (1998a), though the trends over time should be the same. I am not presently aware of the area incorporated in the data used in the above figures, though the strong but not perfect similarity of the time trend between the fall survey plot above and Casey and Myers’ (1998a) one for Georges Bank strongly suggests that that area, but also some others, are included.
The plot of the fall data shows the same pronounced decline in barndoor biomass density in the 1960s, followed by stability, which Casey and Myers (1998a) illustrated for the same areas using much the same dataset. However, as presented here, a very slight biomass increase can be seen in 1993 which was then maintained until 1998. The numbers from the spring surveys (a data set which Casey and Myers seem not to have used) show a similar trend of sharp decline in the late 1960s and early 1970s, followed by stable (if very low) numbers thereafter. The apparent “recovery” of the 1990s, however, began in spring 1992 and has progressed strongly through to 1999. This survey series now indicates (how reliably may be debated) that the barndoor population in U.S. waters, far from sliding into oblivion, is currently at a similar level to that of the late 1960s.
The average weight of the barndoor caught by these surveys is also interesting:
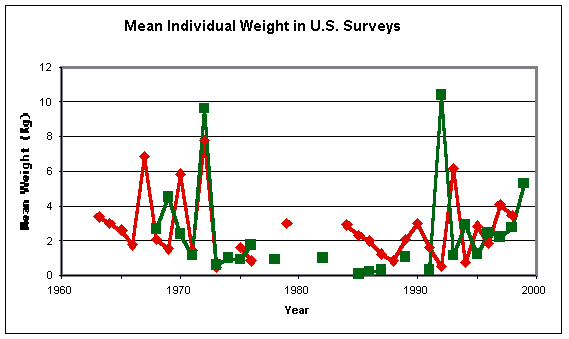
While the data are inevitably confused by the occasional
capture of large individuals, this plot suggests that the biomass decline of
the 1960s was paralleled by a fall in average individual weights, while the
recent “recovery” has seen those weights rise again. This is consistent with
the decline being caused by an increase in mortality rate (perhaps caused by
fishing), which would have initially depleted the older skates more than the
young ones, followed after an interval of some twenty years by the growth of
new year-classes of young barndoor. The same plot is also consistent with some
environmental shift which gradually drove large skates into deep water, beyond
reach of the surveys (or otherwise made them unavailable), but is now allowing
their return. Nevertheless, this is persuasive support for the expected impact
of fishing activity on barndoor abundance.
If the trend has been created by fishing, however, it is notable that the decline occurred under the almost-unrestricted foreign fishing of the 1960s and early 1970s. If that decline was not halted by the international management actions of the last years of ICNAF, the extension of national jurisdiction in 1977 appears to have “cured” the problem. After the expected long lag, during which surviving juvenile barndoor would have had to grow to maturity and spawn in their turn, the population now seems to be trending upwards under national management. Such a scenario may appear to conflict with the very high levels of fishing effort that the U.S. fleet exerted on groundfish in the 1980s and early 1990s. However, that effort was targeted on specific groundfish resources (particularly cod, haddock and yellowtail) and was increasingly focused on hard-bottom areas, where nets with rockhopper or bobbin gear were required. The former eastern European fleets fished different areas, possibly with lighter ground-gear that would have increased their catches of skates. They certainly retained and processed their bycatches of skates and dogfish (Fogarty & Murawski 1998), whereas the U.S. domestic fleet has largely discarded those species, with some unknown proportion of them surviving the experience. Thus, it is by no means improbable that the lower Fishing Mortality rates imposed on groundfish in the 1960s, compared to those of the 1980s and 1990s, resulted in higher rates of incidental Fishing Mortality on barndoor skate.
Such speculations aside, the recent increases in both survey catches-per-tow and individual weights in the survey catches lend strong support to the notion that, far from being in imminent danger of extinction, the barndoor skate is actually undergoing a resurgence – albeit while still at a very low biomass level.
A similarly-brief, preliminary update of the Canadian survey information for NAFO Divisions 4VWX is possible, using data kindly supplied by Marine Fish Division, Bedford Institute of Oceanography. The figures available to me cover the entire area of the three Divisions, rather than the individual ones shown in Casey and Myers, (1998) plots. The result, presented in terms of mean catch per survey tow (rather than biomass per unit area) is:
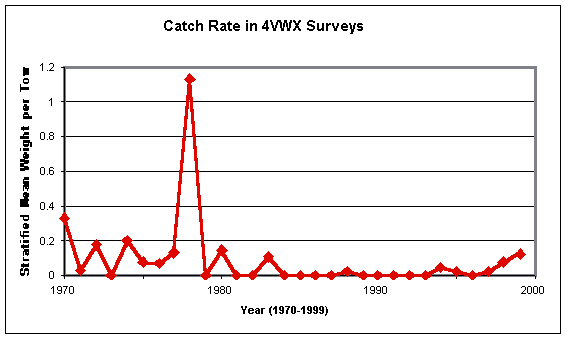
This graph shows the same low values in the 1970s followed by even lower ones in the 1980s as Casey and Myers (1998) found. The single “spike” for 1978 is present in their plots for each of Divisions 4Vs and 4W but is much more prominent when plotted at the scale used here. Inspection of Simon and Comeau’s (1994) map of barndoor catches in these surveys shows that, in 1978, the A.T. Cameron took moderate numbers of the skates on the western side of the Sable Gully and at the head of one of the canyons that cut into the continental slope along the edge of Banquereau. These relatively-good catches were almost certainly nothing more than the chance effects of stratified-random station selection placing the net in prime barndoor habitat. Of more interest is the notable upturn in catches of the species which began in 1996 and has continued to date[20]. While that trend does not appear as dramatic as the one revealed by NMFS’ spring surveys, it is comparable and does lend additional support to the indication of some “recovery” of barndoor that is seen in the U.S. data.
There has been some suggestion that the increased barndoor catches in U.S. waters have been associated with the extensive fishery closures on Georges Bank that were introduced in 1994. There has been no comparable closure on the Scotian Shelf, though bottom fishing was greatly curtailed in Divisions 4VW and reduced in Division 4X in 1993, remaining so to date.
The average weights of the barndoor taken in the Divisions 4VWX surveys point to a larger run of fish on the Scotian Shelf than in U.S. waters, with the averages hovering around 7 kg rather than being mostly below 5 kg:
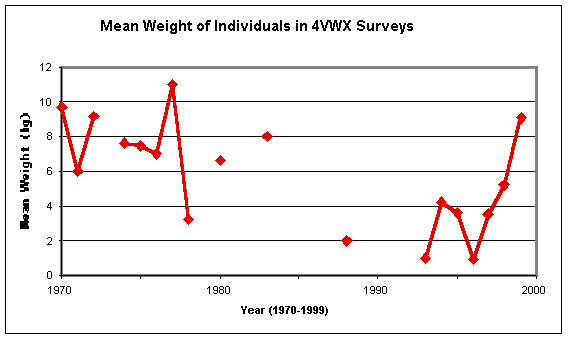
This graph does not really reflect the dip and subsequent rise in barndoor sizes that is evident in the U.S. data, though the Canadian ones could be considered consistent with that trend, if allowance is made for the lack of surveys in the 1960s and the very limited catches through the 1980s.
In addition to these long-established surveys, a new series was initiated in 1999 with the objective of monitoring halibut biomass in deep water. The first such survey evidently encountered significant numbers of barndoor skates (though I have been unable to obtain the relevant data). While the resulting catches cannot indicate biomass trends over time, they may demonstrate the present existence of a substantial “reserve” of these fish below the 365 m-depth limit of both the U.S. surveys and those Canadian ones conducted on the Scotian Shelf. If analysis of the data should lead to that conclusion, it would cast yet further doubt on the notion that the barndoor is in any danger of extinction.
5.1.10 Summary
To summarize this complex of information, it must be understood that the groundfish survey databases do not provide a fully-reliable indication of the biomass of barndoor skate. Since regular standardized surveys commenced (1963 in U.S. waters, 1970 off Nova Scotia, somewhat later off Newfoundland), approximately-comparable data have been collected each year. However, the fishing gear and survey designs used have not been optimized for barndoor and the resulting low catches cannot give convincingly-precise indications of the abundance of that species. Although some data are available for the period before the standardized surveys began, those are not comparable to the later numbers and it is very far from clear that Casey and Myers (1998a) adequately corrected for the many imperfections in the data sets when preparing their plots.
It does appear that the barndoor population was rather stable, though at a low level, from 1970 to 1995, at least in the areas for which Casey and Myers (1998a) compiled the available data. There is some indication that survey catches of the species may have been lower on the eastern Scotian Shelf in the 1980s than it in the 1970s but, even if that decline was real, it could have been an artifact of changed survey gear, rather than a drop in biomass. Preliminary analysis of recent survey data from both the Scotian Shelf and U.S. waters suggests that there has been a significant increase in barndoor abundance since 1995, though it is not impossible that some environmental change has caused a steady biomass to become more available to the survey nets. Clearly, if real, a recent increase in biomass would render untenable any conclusion that the barndoor is endangered.
Before 1970, the surveys did suggest a decline in biomass. There is some solid evidence of the latter stages of this from the U.S. surveys that began in 1963 – though Casey and Myers’ (1998a) inappropriate “correction” for nocturnal fishing will have somewhat exaggerated the extent of the drop. Elsewhere, all that are available are highly dubious data. Casey and Myers’ (1998a) representations of substantial declines in all areas from St. Pierre Bank to NAFO Division 4X through the 1950s and 1960s relied on an undocumented process of selecting which cruises to include, an invalid process for estimating biomass densities from the catches made at stations which did not follow the modern stratified-random survey designs, and the use of data from a variety of ships and nets which will not have yielded comparable catches. Certain of those data were also inappropriately “corrected” for a supposed lack of night-time fishing. The appropriate, rigorous response might be to discard all of the survey information gathered before the advent of the standardized surveys. However, if the barndoor skate did decline markedly, it seems to have done so before 1970 and rejecting the only data which might track that decline is hardly constructive. Thus, I would suggest that there are hints in the available data, especially for St. Pierre Bank, that barndoor did see a marked fall in biomass during the 1950s and 1960s. I would caution, however, that those are only hints, which may well be nothing more than artifacts of the surveys and of Casey and Myers’ (1998a) handling of the data. The “evidence”, if such it is, is by no means as certain as those authors have implied.
Their case might be strengthened by a thorough analysis, by competent fisheries specialists taking full account of the imperfections of the data sets, of the available data from the early surveys and by an examination of just how Casey and Myers (1998a) generated their biomass-density estimates. Such an analysis, along with one that would update the time series to 1998 and 1999, would be valuable but it would require the efforts of both NMFS and Canada’s DFO.
If the decline was real, there is still a question of whether it represented a real fall in the species biomass or only a re-distribution out of the surveyed areas. There are extensive portions of the range of barndoor which Casey and Myers (1998a) ignored, despite survey data being available, those being the Mid-Atlantic Bight and the Gulf of St. Lawrence. There are yet other areas which have not been routinely surveyed, both inshore of the survey strata (which are generally drawn so as to avoid the rocky and complex seabed near the coast) and, more importantly, at great depth along the continental slope. The various surveys have extended well down towards the deepest formally-published catch record for barndoor (various surveys going to 365 or 400 m depth, while the barndoor record is 420 m: Bigelow & Schroeder 1953b). However, Kulka et al.’s (1996) work suggests that the species may live considerably deeper than had been supposed – the limited amount of deep fishing in earlier decades and the limited interest in skates perhaps causing a deficiency in the records.
Supposing that the limited and sketchy data did track a real fall in barndoor biomass in the 1950s and 1960s, there is still a question of whether it was the result of a natural shift in the ocean environment or, alternatively, a consequence of fishing. (There does not seem a reasonable third alternative.) In the St. Pierre Bank region, an environmentally-induced decline is plausible, since the haddock in that area seem to have undergone something of the kind. The limited decline in barndoor biomass in U.S. waters in the 1960s is less easily dismissed. If real, it seems most likely that that was the result of fishing.
As explained above, it is expected that the barndoor should have declined under the pressure of the trawl fisheries which began in the northwest Atlantic early in the 20th Century and expanded greatly in the 1950s, following the invention of the factory-freezer stern trawler. The questions is, perhaps, not whether the barndoor declined but whether the groundfish surveys were able to detect that decline. It seems that they may have measured the last of it in U.S. waters but, further north, only provided vague indications in the years before a standardized survey program was introduced.
The other important question is whether that decline is continuing. The available evidence suggests that it is not. Following 25 years of near-stability, perhaps with a slow dip to a low point during the 1980s, the barndoor population is now showing some signs of “recovery”. The timing of the onset of near-stability and that of the commencement of biomass increase suggest that the modern U.S. and Canadian fisheries were not the cause of the earlier decline. Rather, the barndoor seems, in so far as anything is known, to have stood up well to the extreme fishing efforts being applied to groundfish in the later 1980s and early 1990s. Instead, it seems to have declined (if it declined at all) during the period of the foreign fisheries from the early 1950s up to the mid-1970s. The modern fisheries may be too closely targeted on their intended resources to take many barndoor and may use gear (particularly with bobbins and rockhoppers) which roll over skates without harming them. The former foreign fisheries, in contrast, fished more widely (lacking the sophisticated electronics now used) and may have often used nets with simple groundropes. As such, they would have caused higher bycatch moralities for the same effective effort on their target species.
Whatever the actual cause of the earlier decline (and even supposing that it did occur as it should have been expected to do), there is no sign that modern fishing is having an adverse effect, nor that barndoor are experiencing a continuing decline. Quite the reverse.
5.2 Critical Mortality Rates
5.2.1 Vulnerability of Barndoor to Fishing Gear
Casey and Myers (1998a) noted that barndoor skate are vulnerable to trawling from their time of hatching onwards, unlike the target species of the fishing (cod, flounder etc.) which are able to pass through the meshes until they have grown to about their sizes at sexual maturity. That is an important and valid point. However, to say that the barndoor are vulnerable to trawls throughout their lives is not to say that they are fully vulnerable.
Casey and Myers’ (1998a) brief comment on the point relates only to escapement through the meshes: Once a barndoor of almost any size is within a trawl net, it will be unable to escape since even the recently-hatched ones would have trouble passing through the webbing of a groundfish trawl. There are, however, two stages to a skate’s vulnerability to trawling before it finds itself within the net. Firstly, commercial fishing activity is not uniformly spread across the ocean floor but is concentrated in areas which promise reasonable catches of the target species. Barndoor is not a target of any fishery, nor has it ever been. Its geographical and depth ranges are not identical to those of any of the targets – particularly since the skate seem, on average, to live at greater depths than most commercial species of the northwest Atlantic (save halibut, tilefish, redfish and grenadiers, of which only the latter two have supported significant trawl fisheries). Moreover, while the habitat preferences of the various species are not well known (and those of barndoor are almost unknown), it is exceedingly unlikely that barndoor skate choose to aggregate in quite the same places as cod, haddock, yellowtail or any of the other target species do. That is not to say that barndoor are never taken as bycatch. Clearly they are. However, a cod fishery which took 33% of the Georges Bank cod in a year (imposing a Fishing Mortality rate of 0.4), with nets optimized for cod and so capable of catching most of those in their path, would encounter far less than 33% of the barndoor skate on and around the Bank during that fishing. How much less cannot be estimated (depending as it does, in part, on how different the micro-scale distributions of the species are and on how well the fishermen can target the type that they are after), though the “availability” of barndoor to the nets could easily be ten times less than those of their target species.
Once a trawl is deployed with a skate in its path, there is still a question of whether the animal can evade the net before entering its mouth. Unlike large sharks, which probably out-swim the gear, or cod, which try to rise above the headrope, skate seem to hung the bottom and let the net run over them. As already noted, limited comparative fishing studies suggest that this behaviour results in catches with bobbin gear of five to ten times less than those in nets with simple footropes. Even the latter may fail to take some skates in their paths, however. Thus, if only one tenth of the barndoor lay in the path of the nets of a hypothetical fishery taking 33% of the cod, perhaps only one tenth of those would be taken, resulting in a barndoor bycatch of 0.3% of the population.
While these “estimates” are clearly highly uncertain, they do illustrate the key point: The undoubted vulnerability of all sizes of barndoor skate to retention inside trawl nets does not equate to this bycatch species being anywhere near as vulnerable to trawling as adult fish of the target species are.
There is also a question of the fates of such barndoor as are caught. Bony fish typically have gas-filled swimbladders, which expand as the fish is hauled to the surface, often causing fatal injuries. Skates, however, have no gas-filled cavities. Moreover, like their “cousins” the dogfish, they give every appearance once on board of being “tough” and resilient to damage. While there seems never to have been a study of how well skate survive being caught and discarded, there is every possibility that some considerable proportion of them is returned to the sea unharmed (cf. Murawski 1996)[21].
5.2.2 Mortality Rate Model
When he first raised the plight of the large skates, Brander (1981) advanced a simple model of the population dynamics of Raja batis in which he equated the death rate of mature fish to the recruitment rate of young adults. Clearly, those two must be equal if the population is to remain stable, while the latter can be estimated from the annual egg production per female and the death rate of the juveniles. Extrapolation from the fecundities of other skates suggested that R. batis would spawn some 40 eggs per female per year. From previous research on the species, the age at maturity was taken to be 11 years. Brander (1981) was then able to draw a graph showing the combinations of juvenile and adult death rates that would produce a stable population. (High death rates imposed on the adults would be sustainable so long as the death rate of the juveniles was low and vice versa.) If the two death rates were identical, Brander’s (1981) calculations indicated that both would have to be 0.37 (31% dying each year) for the population to remain stable[22].
In so far as it went, Brander’s (1981) model was straightforward and not objectionable. However, from it he drew the inference that death rates higher than indicated by his graph would “result in the collapse of the population”. That was wrong. All populations always have some death rate which, if it could be achieved, would maintain their stability. However, since stability never actually occurs in our variable world, the dynamics of each population contain some “control mechanism” which allows increased production when abundance falls and cuts production when numbers increase, thus (usually) avoiding either extinction or over-population. Were it not so, the biosphere would not contain the great diversity of approximately-stable species that it does.
Brander’s (1981) calculations actually found, not the death rates that would cause a collapse of the population, but rather those that would produce stability under the conditions which cause Raja batis to spawn 40 eggs per female per year and to produce growth to sexual maturity in eleven years. Exceeding those mortality rates would certainly cause the population to fall but that would in turn lead to some increase in growth rates, generating younger ages at maturity and higher effective fecundities – thus restoring stability. To produce a “collapse” it would be necessary to exceed, not the death rate for stability at the current level, but the species’ ability to tolerate higher death rates at lower population levels. That ability is poorly known for any fish and entirely unknown for barndoor or R. batis. Their “resilience”, if it may be so called, will be low when compared to those of most bony fish (which are far more fecund) but it will exist. Brander’s (1981) model was thus either irrelevant to the issue which concerned him or, at best, provided a minimum estimate of the critical death rate which he sought – the death rate that would produce a collapse of the population.
Casey and Myers (1998a) repeated Brander’s (1981) logic, including its error, which they extended yet further by declaring that death rates higher than those calculated to produce stability would “drive the barndoor skate to extinction”. The more-recent authors used a different mathematical formulation but one that was functionally identical to Brander’s (1981)[23]. They followed his estimate of the age at maturity (11 years), deeming that sufficiently accurate when applied to barndoor skate, and estimated fecundity at 47 eggs per female per year using the same extrapolation as Brander (1981) had applied to his species. That led them to suggest that the critical death rate was around 0.4, though they also advanced the notion that growth rates would be lower at the northern limit of the species’ range (meaning Grand and St. Pierre banks) and so that the mortality rate of concern would drop to 0.2 in some areas. This last was an entirely groundless assertion, perhaps based on a dubious extrapolation from the dynamics of the various cod populations in the northwest Atlantic.
5.2.3 Bycatch Mortality Rates Imposed by the Fisheries
Casey and Myers (1998a) did not spell out why they though the barndoor skate was approaching extinction (as distinct from simply being depleted) nor did they directly state which human agency they supposed was responsible. However, they did suggest that an extensive closure to the trawl fisheries was needed as “[p]erhaps the only hope” for the species and their paper certainly implies that they supposed that those fisheries in particular were responsible for the perceived situation.
They also correctly noted that the Fishing Mortality rates exerted on northwest Atlantic cod have long been greater than the 0.4 which they supposed was critical for the survival of barndoor skate (Casey & Myers 1998a). In so far as can be judged (and recognizing that stock assessments are not highly precise calculations), that rate has indeed been above 0.4 in most areas since some time in the 1960s. Since cod also suffer a Natural Mortality rate that is conventionally assumed to be 0.2, their Total Mortality rate [equivalent to the rates calculated for skates by Brander (1981) and Casey and Myers (1998a)] has generally been above 0.6. Thus, it seems safe to conclude that all cod populations in the region have experienced death rates well above 0.4 for many years.
Having persuaded themselves that barndoor are vulnerable to trawl nets, and that they have a critical death rate, beyond which they will be driven to extinction, of between 0.2 and 0.4, it is perhaps not surprizing that Casey and Myers (1998a) were led to the conclusion that the species was bound for extinction. As shown above, however, those death rates relate to stability, not extinction, while the “vulnerability” of barndoor to trawling directed for other species is very much less than 100% – perhaps a hundred times less. In short, Casey and Myers (1998a) advanced no valid grounds whatsoever for supposing that the trawl fisheries could drive the barndoor to extinction.
It is to be expected that fishing has led to the depletion of barndoor and, as shown above, there is some indication in the survey data that that has indeed happened. However, biological extinction of such an open-sea species has not yet happened as a result of fishing alone. Nor is there any reason to suppose that the barndoor is vulnerable to such a fate.
5.3 Extinction of Raja
batis
Casey and Myers’ (1998a) statement that Raja batis had been shown to be locally extinct in the Irish Sea was supported only by a citation of Brander’s (1981) paper. Brander himself made no such claim in that work and indeed stated in his abstract that his species was “now very rare”. He did document a major reduction in its abundance since 1902, which doubtless constituted severe depletion. He also noted that no R. batis had been caught by government research vessels in the Irish Sea in the previous ten years (presumably meaning the 1970s), while the only specimens landed in Fleetwood by commercial vessels in the five years before he wrote had been caught in the North Channel (between Ireland and Scotland), rather than within the Irish Sea itself. He was, however, careful to use such terms as “disappearance”, never “extinction”, to describe the state of the skates (Brander 1981).
It should also be noted that the range of Raja batis extends from northern Russia and Iceland to Algeria, within which vast area it reaches depths as great as those inhabited in the northwest Atlantic by barndoor skate. Even if the species were entirely extirpated from the Irish Sea, that would still be very far from representing biological extinction of such a wide-ranging species. Thus, while the sorry tale of the (now-misnamed) “common skate” is another demonstration of the vulnerability of such animals to severe depletion, it gives no support whatever to the improbable notion that human actions can lead to the biological extinction of a fish species of the open sea.
5.4 Refuges and Closures
In addition to presenting their case for barndoor skate being near extinction, Casey and Myers (1998a) made some comments on fishery closures. Specifically, they suggested that the continued “survival” (meaning occurrence in research-vessel survey catches) of barndoor on “Browns Bank” (meaning in NAFO Division 4X) and on Georges could probably be accounted for by the higher temperatures in those areas than further north, which would allow faster growth rates and younger ages at maturity, and also to the seasonal closures of parts of those banks to “trawling”. The closures in question are not, in fact, to the trawl fisheries but rather to all groundfish fishing, though there formerly were exemptions in some of the areas on Georges Bank which allowed hook-and-line fishing provided that only large hooks, which take few haddock, were used (Halliday 1988).
I find the suggestion that these closures had a significant influence on the present status of barndoor to be highly improbable. Firstly, while the closures may have somewhat reduced the fishing effort exerted on cod, haddock and yellowtail relative to what it might otherwise have been, they did not serve to keep the Fishing Mortality exerted on those resources any lower than the intensities exerted on the eastern Scotian Shelf or St. Pierre Bank – areas without such closures and where barndoor disappeared from the survey catches. The fishing effort exerted in Divisions 4X and 5Y was forced into open areas and open seasons but it was not diminished by the closures. Thus, unless the overlaps in space and time which placed skate in the track of gear set for groundfish were particularly concentrated in the areas and seasons of the closure (a highly improbable contention), barndoor should have experienced as much bycatch mortality in the groundfish fisheries in Divisions 4X and 5Y as they did anywhere else.
Secondly, for the closures to have had the effect suggested by Casey and Myers (1998a), it would be (almost) essential that they covered an appreciable part of each of the two Divisions. In reality, the Georges closures covered very much less than the entire Bank, while that on Browns enclosed only some 20% of Division 4X. Indeed, most of the barndoor skate taken by the research-vessel surveys in that Division (Simon & Comeau 1994) were caught outside the boundaries of the closed area (Halliday 1988).
Casey and Myers (1998a), however, proceeded with their opinion that these seasonal closures had had a significant role in the local “survival” of barndoor and thus concluded that “[p]erhaps the only hope for the survival of this species is to designate an area protected from trawling on all the banks that is sufficiently large to allow for a self-sustaining population”. While such a closure would doubtless advance the agendas of various political interests, there is no reason to suppose that the survival of barndoor is in any danger, nor is there any particular reason to suppose that such a closure would have a marked effect on the population. Indeed, considering that barndoor “bites readily on almost any bait, and is often caught on hand and long lines as well as in otter trawls” (Bigelow & Schroeder 1953a), there can be no scientific foundation for recommending a trawling closure, as distinct from a closure covering all groundfish fishing, as a measure to protect this species.
6 Considerations of Endangerment
Species may be listed under the Endangered Species Act on a number of grounds. This document cannot address the legalities of such a listing but a few points can be made from a scientific perspective.
6.1 Utilization
Barndoor skate seem never to have been a target (not even a secondary species of interest) of any fishery, though for over a hundred years (e.g. Goode 1884) some of those caught incidentally have been utilised by mankind. The incidental catch may sometimes have been considerable and sufficient to deplete the populations of barndoor, given their potential vulnerability to the effects of fishing. However, as explained above, such bycatch mortality in fisheries directed for other species has not and will not be intensive enough to lead to biological extinction of barndoor.
6.2 Regulatory Regimes
There is no regulatory regime specifically designed to protect barndoor skate. However, the fisheries in which it is incidentally taken are all subject to regulation. The management systems in both the United States and Canada failed to maintain the target resources at optimal levels during the 1980s and early 1990s but the improved and strengthened arrangements now in place are slowly leading to some re-building of over-fished resources. Since the intensive fishing of the past did not eliminate the barndoor, there is no reason to suppose that the regulations made under the Sustainable Fisheries Act will prove insufficient for its protection, despite that not being their intended purpose.
There may be some need to pay special attention to bycatch issues if new fisheries are developed. Deepwater fishing for tilefish and other species, for example, might impose new pressures on barndoor, requiring a degree of consideration beyond those normally given to incidental catches. Even that possibility, however, has already been (inadvertently) anticipated in the Sustainable Fisheries Act’s provisions relating to bycatches in general.
6.3 Factors Affecting Abundance
Aside from the known and expected depletion effect caused by fishing, there is little evidence that the abundance of barndoor skate is subject to any particular pressures. They are, of course, affected by the ocean environment and its natural changes (some of which may be indirectly influenced by mankind through global warning). As noted above, there is some indication in the field data that barndoor has withdrawn from shallower areas at the northern part of its range in response to some unknown environmental shift. Such range extensions and contractions are to be expected in marine populations.
The effects of anthropogenic pollution and coastal-zone development, while doubtless serious in some inshore waters where barndoor once occurred, do not extend outwards (at significant levels) to the offshore banks and continental slopes which represent the core of the species’ range. Indeed, the current pristine state of the waters on Georges Bank was a major factor in the recent recommendation to maintain a moratorium on oil and gas activity in the Canadian portion of that area.
6.4 Disease and Predation
There is little reason to suppose that barndoor are subject to any unusual biological pressures. The only known factor which might conceivably have some influence would be increased predation by the burgeoning population of grey seals on the eastern Scotian Shelf, which may now be of sufficient magnitude to affect the cod population in that area (Mohn & Bowen 1996). Given the size, depth distribution and behaviour of barndoor, however, it seems grossly improbable that the seals could have a significant impact. They are, in any event, only abundant over a small proportion of the range of the skate.
6.5 Threats to Habitat
The offshore habitat of the barndoor is almost immune to human threat. There is some possibility of habitat modification by the offshore petroleum industry operating off Nova Scotia and Newfoundland which may require close attention during future environmental assessments. However, there is no reason to suppose that the level of habitat damage would be such as to eliminate a species with the broad range of tolerances shown by barndoor.
It is widely supposed that some fishing gear, particularly scallop rakes and otter trawls, is modifying hard-bottom communities that may constitute important habitat for some fish, such as the Georges Bank cod. Barndoor skate, however, show no preference for rocky areas and indeed seem commonest now on the muddy bottoms of the continental slope. Their habitat, therefore, is not such as to be threatened by fishing gear.
7 Conclusions
In conclusion, it was to be expected that barndoor skate would be depleted by the bycatch mortality imposed on them by fisheries for other species. The available survey data are not unequivocal but do seem to track such depletion in the years before 1970, though some of the apparent decline may have natural, environmental causes, rather than being linked to fishing. Those data certainly do not support the picture of extreme declines presented by Casey and Myers (1998a), which was exaggerated and relied on highly-uncertain interpretations of dubious data. From 1970 to 1995, barndoor abundance seems to have stabilized, in so far as anything can be judged from the available data. There may have been some on-going decline into the 1980s but that appears to have been more than offset by a modest recent (post-1995) increase.
There is no solid evidence, nor even vague indications, that the supposed decline in the barndoor before 1970 was caused by the expected effects of fishing, though it would be appropriately precautionary to assume that it was. Casey and Myers’ (1998a) suggestion that the bycatch mortality, exerted on barndoor by fisheries for other species, is sufficient to lead to their extinction was fallacious, being founded on an inappropriate model of skate dynamics and a misunderstanding of the fisheries concerned. There is no reason to suppose that the death rates imposed by the fishery were ever sufficient to drive this species to extinction, nor ever likely will be. Moreover, the recent increase in survey catches of the species suggests that the fishery of the 1990s is exerting less pressure on the species than it bore in the years of international fishing. The serious situation once facing the barndoor appears to have been resolved twenty years before it attracted the attention of environmentalists.
When he first raised the loss of Raja batis from the Irish Sea, Brander (1981) remarked that its disappearance was not surprizing but the fact that its loss had passed unnoticed and without comment was. Casey and Myers (1998a) took up that theme, suggesting that the failure to examine historical data had resulted in the largest skate in the northwest Atlantic being driven to “near extinction” without anyone noticing. They even hinted that other, “little known” species may face a worse fate[24]. They also suggested that it is necessary to assemble long-term and wide-area data so as to detect such “near extinctions”. In so far as it went, that recommendation was sensible, though one might ask why they did not apply their own advice by studying the whole range of the species of concern, along with the broader contemporaneous events affecting others, such as haddock. It was not, in nay case, entirely fair to claim, as Casey and Myers (1998a) did, that the decline in barndoor had passed “without anyone noticing”. Grosslein (1972), for one, remarked on the 50% decline in skate biomass on Georges Bank shown during the first decade of regular U.S. groundfish surveys – a trend he correctly regarded as “not at all surprizing in view of the tremendous increase in bottom trawling activity and the relatively low reproductive potential of these species”. What was lacking was not so much recognition of the depletion of barndoor but rather, before the 1990s, any public or policy concern over the status of non-commercial fish.
Finally, Casey and Myers (1998a) call for more attention to long-term data must carry a caveat. Except in the cases of the few rigorously-standardized monitoring programs that have been continued over very long periods, reaching back more than a few decades requires the careful use of data gathered by a complex of non-comparable methods. Interpreting such data needs a detailed and extensive understanding of the means by which it was collected and of the system being monitored. The sort of naïve, uncritical and poorly-informed compilation of old data sets to which Casey and Myers (1998a) resorted will generally lead to erroneous conclusions – as it did in the case of their study of barndoor skate. In this document, I have attempted to supply some of the missing understanding and critical attention. After doing so, I can draw only one conclusion from the available evidence: The barndoor skate is not near to biological extinction and is showing no sign that it is headed in that direction. Rather, it seems to be experiencing a slow increase in abundance, in a setting where human activity poses no threat to its continued existence as a species.
Cited References
Auster, P.J., R.J. Malatesta & S.C. LaRosa (1995) Patterns of microhabitat utilization by mobile megafauna on the southern New England (USA) continental shelf and slope. Marine Ecology Progress Series 127: 77-85.
Azarovitz, T.R. (1981) A brief historical review of the Woods Hole Laboratory trawl survey time series. In: W.G. Doubleday & D. Rivard (eds.) Bottom Trawl Surveys: Proceedings of a Workshop held at Ottawa, November 12-14, 1980. Canadian Special Publication of Fisheries and Aquatic Sciences 58: 62-67.
Bell, R. (1859) On the natural history of the Gulf of St. Lawrence, and the distribution of the mollusca of eastern Canada. Canadian Naturalist and Geologist 4: 197-220.
Bigelow, H.B. & W.C. Schroeder (1953a) Fishes of the Gulf of Maine. Fishery Bulletin, U.S. Fish & Wildlife Service 74(53).
Bigelow, H.B. & W.C. Schroeder (1953b) Fishes of the Western North Atlantic: Part Two: Sawfishes, Guitarfishes, Skates and Rays. Sears Foundation for Marine Research, Memoir Number 1.
Brander, K. (1981) Disappearance of common skate Raia batis from Irish Sea. Nature 290: 48-49.
Casey, J.M. & R.A. Myers (1998a) Near extinction of a large, widely distributed fish. Science 281: 690-692.
Casey, J.M. &
R.A. Myers (1998b) Diel variation in trawl catchability: is it as clear as
day and night? Canadian Journal of
Fisheries and Aquatic Sciences 55: 2329-2340.
Cornish, G.A. (1912) Notes on the fishes of Tignish, Prince Edward Island. Contributions to Canadian Biology 1906-10: 79-81.
Cox, P. (1921) List of fishes collected in 1917 off the Cape Breton coast and the Magdalen Islands. Contributions to Canadian Biology 1918-20: 109-114.
Fogarty, M.J. & S.A. Murawski (1998) Large-scale disturbance and the structure of marine systems: Fishery impacts on Georges Bank. Ecological Applications 8(1 Supplement): S6-S22.
Goode, G.B. (1884) The Fisheries and Fishery Industries of the United States: Section I: Natural history of useful aquatic animals. Government Printing Office, Washington.
Griswold, B.L. & V. Efanov (1972) USA-USSR Cooperative Fishery Research in 1971. ICNAF Res.Doc. 72/112, Serial No. 2838.
Grosslein, M.D. (1968) Results of the joint USA-USSR groundfish studies. Part II: Groundfish survey from Cape Hatteras to Cape Cod. ICNAF Res.Doc. 68/87, Serial No. 2075.
Grosslein, M.D. (1972) A preliminary evaluation of the effects of fishing on the total fish biomass, and first approximations of Maximum Sustainable Yield for finfishes, in ICNAF Division 5Z and Subarea 6. Part 1. Changes in relative biomass of groundfish in Division 5Z as indicated by research vessel surveys , and probable maximum yield of total groundfish resource. ICNAF Res.Doc. 72/119, Serial No. 2835.
Halliday, R.G. (1988) Use of seasonal spawning area closures in the management of haddock fisheries in the northwest Atlantic. NAFO Scientific Council Studies 12: 27-36.
Halliday, R.G. & P.A. Koeller (1981) A history of Canadian groundfish trawling surveys and data usage in ICNAF Divisions 4TVWX. In: W.G. Doubleday & D. Rivard (eds.) Bottom Trawl Surveys: Proceedings of a Workshop held at Ottawa, November 12-14, 1980. Canadian Special Publication of Fisheries and Aquatic Sciences 58: 27-41.
Hennemuth, R.C. (1968) Results of the joint USA-USSR groundfish studies. Part I: Comparative fishing experiments. ICNAF Res.Doc. 68/86, Serial No. 2074.
Huntsman, A.G., W.B. Bailey & H.B. Hachey (1954) The general oceanography of the Strait of Belle Isle. Journal of the Fisheries Research Board of Canada 11: 198-260
Kenchington, T.J. (1980) The fishes of St. Georges Bay, Nova Scotia. Canadian Technical Report of Fisheries and Aquatic Sciences 955: 154 p.
Kenchington, T.J. (1996) Long-term stability and change in the commercial groundfish longline fishing grounds of the northwest Atlantic. Fisheries Research 25: 139-154.
Kenchington, T.J.
& R.G. Halliday (1994) A survey of fishing practices in the Scotia-Fundy
Region groundfish longline fisheries. Canadian
Manuscript Report of Fisheries and Aquatic Sciences 2225: 642 p.
Kulka, D.W., E.M. DeBlois & D.B. Atkinson (1996) Non-traditional groundfish species on Labrador Shelf and Grand Banks – Skate. DFO Atlantic Fisheries Research Document 96/98.
Leim A.H. & W.B. Scott (1966) Fishes of the Atlantic coast of Canada. Bulletin of the Fisheries Research Board of Canada 155.
McEachran, J.D. & J.A. Musick (1975) Distribution and relative abundance of seven species of skates (Pisces: Rajidae) which occur between Nova Scotia and Cape Hatteras. U.S. Fishery Bulletin 73: 110-136.
McKenzie, R.A. (1946) The haddock fishery of grounds fished by Canadians. Bulletin of the Fisheries Research Board of Canada 69: 30p.
McKenzie, R.A. (1959) Marine and freshwater fishes of the Miramichi River and estuary, New Brunswick. Journal of the Fisheries Research Board of Canada 16: 807-833.
Merriman, D. & H.E. Warfel (1944) Studies of the marine resources of southern New England – II. A preliminary analysis of the Connecticut trawl fishery. Transactions of the Ninth North American Wildlife Conference: 230-239.
Mohn, R. & W.D. Bowen (1996) Grey seal predation on the eastern Scotian Shelf: modelling the impact on Atlantic cod. Canadian Journal of Fisheries and Aquatic Sciences 53: 2722-2738.
Murawski, S.A. (1996) Biological implications of bycatch. In: K. Castro, T. Corey, J. deAlteris & C. Gagnon (eds.) Proceedings of the East Coast Bycatch Conference. Rhode Island Sea Grant: 31-39.
Needler, A.B. (1931) The haddock. Bulletin of the Biological Board of Canada 25: 28p.
Neilson, J.D.,
K.G. Waiwood & S.J. Smith (1989) Survival of Atlantic halibut (Hippoglossus hippoglossus) caught by
longline and otter trawl gear. Canadian
Journal of Fisheries and Aquatic Sciences 46: 887-897.
Pitt, T.K., R. Wells & W.D. McKone (1981) A critique of research vessel otter trawl surveys by the St. John’s Research and Resource Services. In: W.G. Doubleday & D. Rivard (eds.) Bottom Trawl Surveys: Proceedings of a Workshop held at Ottawa, November 12-14, 1980. Canadian Special Publication of Fisheries and Aquatic Sciences 58: 42-61.
Raloff, J. (1999) Skate-ing to extinction? Some long-lived fish are facing accidental annihilation. Science News 155: 280-282.
Scott, J.S. (1977) Groundfish landings and movements related to the Strait of Canso area. Fisheries and Marine Service Technical Report 834(3): 1-7.
Scott W.B.
& M.G. Scott (1988) Atlantic fishes of Canada. Canadian Bulletin of Fisheries and Aquatic Sciences 219: xxx+731p.
Simon, J.E. & P.A. Comeau
(1994) Summer distribution and abundance trends of species caught on the
Scotian Shelf from 1970-92, by the research vessel groundfish survey. Canadian Technical Report of Fisheries and
Aquatic Sciences 1953.
Vladykov, V.D. (1936) Capsules d’oeufs de raies de l’Atlantique Canadien appartenant su genre Raja. Le Naturaliste Canadien 63: 211-231.